Zinc and Regulation of Inflammatory Cytokines: Implications for Cardiometabolic Disease

Abstract
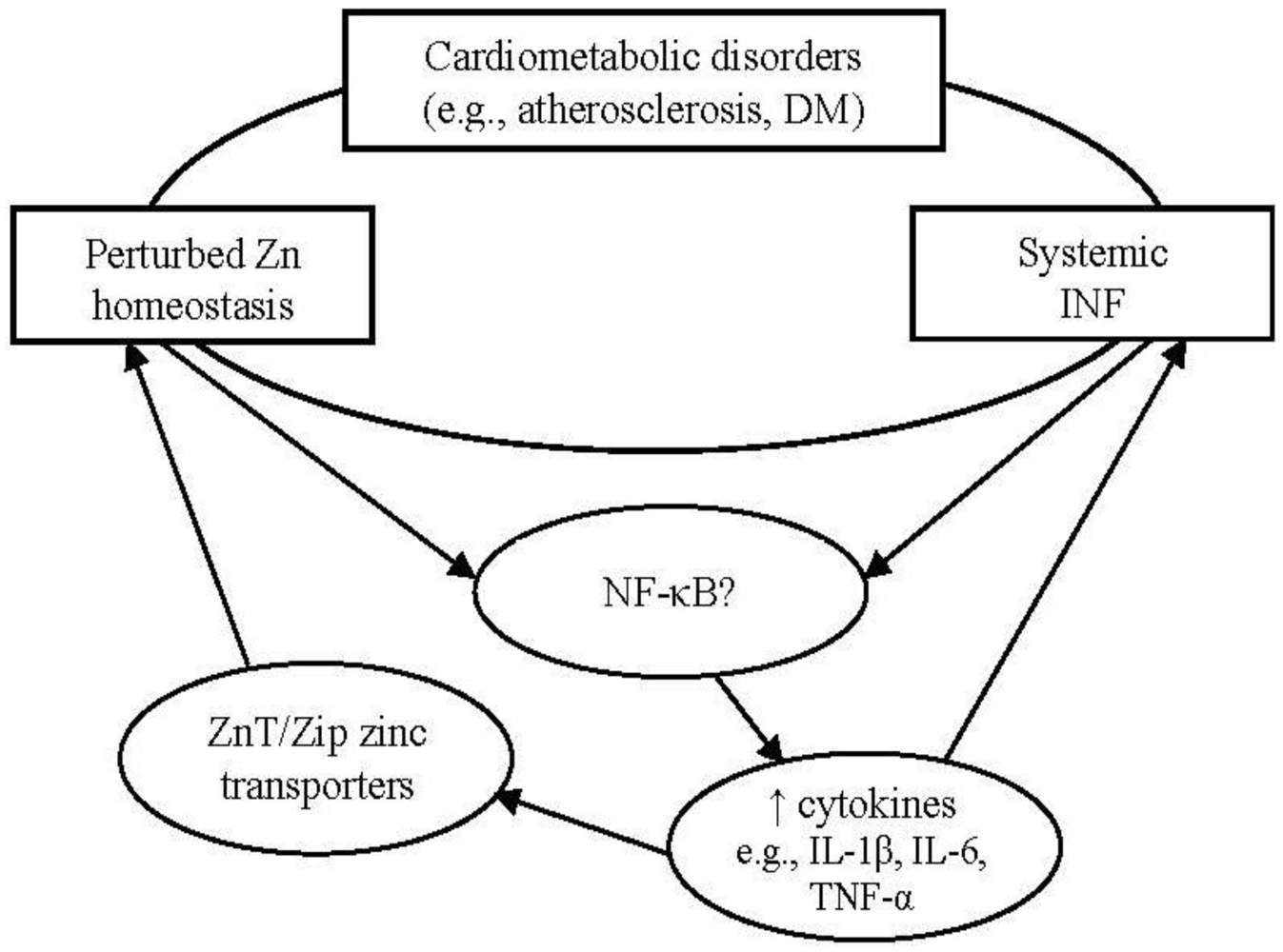
:1. Introduction

{kind=link}
| Cytokine | Primary Cell Sources | Key Functions in Inflammation |
|---|---|---|
| IL-1 | Macrophages Endothelial cells | Synthesis of acute phase proteins by hepatocytes; Local and systemic inflammatory effects |
| IL-2 | Activated T cells Th1 cells | Proliferation of T cells, B cells; Proliferation and activation of NK cells |
| IL-6 | Macrophages Th2 cells Endothelial cells Adipocytes Myocytes Osteoblasts | Synthesis of acute phase proteins by hepatocytes; Proliferation of B cells; Down-regulation of IL-1 and TNF production; Activation of immune cells, osteoclasts, endothelial cells; Hypothalamic Pituitary Axis—fever & hormone release |
| IL-10 | MacrophagesTh2 cells | Resolution of inflammation; Inhibition of Th1 inflammatory cytokine synthesis; Inhibition of activated macrophages and dendritic cells |
| IL-12 | Macrophages Dendritic cells | Promotion of Th1 differentiation; Stimulation of IFN-γ production by T cells, NK cells |
| TNF-α | Macrophages T cells NK cells Lymphoid cells Endothelial cells Adipocytes Cardiac myocytes Fibroblasts Neuronal cells | Synthesis of acute phase proteins by hepatocytes; Recruitment and activation of neutrophils and monocytes at sites of infection; Stimulation of CRP release from liver; Activation of NF-κB pathway; Induction of insulin resistance |
| TGF-β | Macrophages T cells | Resolution of inflammation; Limit production of IL-2, IFN-γ, and TNF; Inhibition of proliferation/activation of B cells, T cells, macrophages. |
| IFN-γ | Th1 cells NK cells | Activation of macrophages; Suppression of Th2 cell activity; Promotion of leukocyte migration |
2. Zinc Homeostasis and Inflammation
3. Zinc Status and Cytokines
3.1. Cytokines in Chronic Inflammation
3.2. Human Zinc Deficiency
3.3. Zinc Supplementation Studies in Humans
3.4. In Vitro Studies in Human Cells
| Author, Year | Description of Participants | Study Design/Cell Culture Conditions | Zn Dose (mg/day) | Outcome |
|---|---|---|---|---|
| Bao et al., 2008 [34] | 36 adults with sickle cell disease; M & F; 18–47 years | RCT, parallel, 13 weeks; isolated PBMCs stimulated with LPS or PHA-P for 24 h | 75 | Decrease in LPS-induced TNF-α & IL-1β mRNA & protein concentrations & increase in PHA-P induced IL-2 mRNA concentrations in Zn compared to placebo group |
| Prasad et al., 2007 [35] | 50 healthy older adults; M & F; 55–87 years/whole blood stimulated for 4 h & isolated PBMCs for 24 h with LPS | RCT, parallel, 52 weeks; isolated whole blood cells or PBMCs stimulated with LPS for 4 or 24 h | 45 | No change in % of whole blood cells positive for IL-1β or TNF-α in Zn supplement group; decrease in ex vivo generation of TNF-α in PBMCs from Zn supplement group |
| Bao et al., 2010 [36] | 40 healthy older adults; M & F; 56–83 years | RCT, parallel, 26 weeks | 45 | Decrease in plasma IL-6 concentrations in Zn supplement group |
| Prasad et al., 2004 [37] | 20 healthy adults; M & F; 19–50 years | RCT, parallel, 8 weeks; isolated PBMCs stimulated with LPS for 24 h | 45 | Zn reduced concentrations of LPS-induced TNF-α & IL-1β mRNAs |
| Raqib et al., 2004 [38] | 56 Shigella-infected children; M & F; 1–5 years | RCT, parallel, 2 weeks; isolated PBMCs stimulated with PHA for 72 h | 20 | No significant effects of Zn on PHA-induced release of IL-1β, IL-2, or IFN-γ |
| Sandstead et al., 2008 [39] | 54 children; M & F; 6–7 years | RCT, parallel, 10 weeks; isolated PBMC stimulated with PHA-P for 48 h | 20 a | Greater release of IL-2 & IFN-γ in stimulated cells derived from Zn supplemented compared to control subjects |
| Aydemir et al., 2006 [40] | adults; M; 19–31 years | controlled before & after trial, 10 days; isolated cells stimulated with LPS for 2 h or by antigen presentation for 2 days | 15 | Greater release of TNF-α, IL-1β, IFN-γ in stimulated cells (monocytes, granulocytes, & T lymphocytes, respectively) derived from Zn supplemented subjects compared to placebo |
| Kahmann et al., 2008 [41] | 19 healthy older adults; M & F; 65–82 years | uncontrolled before & after trial, 48 days; isolated PBMCs stimulated with LPS or SPEA for 72 h | 10 | Zn supplementation resulted in lower basal IL-6, higher LPS-induced IL-6, & higher SPEA-induced TNF-α & IFN-γ concentrations |
| Mariani et al., 2008 [42] | 39 healthy older adults; M & F; 60–83 years | uncontrolled before & after trial, 48 days | 10 | Increase in plasma IL-6 concentrations with Zn |
| Mocchegiani et al., 2008 [43] | 110 healthy older adults; M & F; 65–85 years | uncontrolled before & after trial, 48 days | 10 | No change in TNF-α but increase in plasma IL-6 concentrations with Zn |
| Kara et al., 2011 [44] | 40 athletes and sedentary young adults; M; 15–17 years | controlled before & after trial, 8 weeks | 5 b | Zn supplementation resulted in higher serum IL-2, TNF-α & IFN-γ concentrations compared to non-supplemented individuals, irrespective of exercise |
| Author, Year | Treatment | Outcome |
|---|---|---|
| Metz et al., 2007 [50] | PBMC were supplemented with 30, 60 µM Zn or 1 µM TPEN for 7 days before stimulation with PHA for 24 h | PHA-stimulated IFN-γ & IL-10 concentrations were higher in cells pre-treated with 60 µM Zn |
| Poleganov et al., 2007 [51] | PBMC were supplemented with 7.5, 15, 30 µM Zn & stimulated with IL-1β, IL-12, or IL-18 for 39 h | Zn amplified the induction of IFN-γ by IL-1β, IL-12 & IL-18 |
| Chang et al., 2006 [52] | PBMC were treated with 3, 30, 100, 300, 1000 µM Zn for 24 h | TNF-α, IL-1β, IL-6, IL-12, and IFN-γ concentrations increased at zinc concentrations ≥100 µM in combination with decreased cell proliferation |
| von Bulow et al., 2005 [53] | PBMC were treated with 25 µM Zn plus 50 µM pyrithione & stimulated with LPS for 24 h; primary monocytes were incubated with 25, 125 µM Zn for 1 h before addition of LPS (250 ng/mL) for 24 h, or stimulated with 1 µM Zn plus pyrithione (50 µM) & LPS for 24 h | Application of zinc plus pyrithione blocked LPS-induced release of IL-1β & TNF-α in PBMC; in monocytes, 125 µM Zn significantly inhibited TNF-α release compared to controls; the application of zinc plus the pyrithione ionophore abrogated LPS-induced release of IL-1β & TNF-α |
| Wellinghausen et al., 1997 [54] | PBMC in serum-free medium were stimulated with 100 µM Zn for 48 h | Zn increased IL-1β concentrations; Zn-induced secretion of IFN-γ was not measurable |
| Driessen et al., 1995 [55] | PBMC were supplemented with 12.5, 25, 50, 100 µM Zn & stimulated with LPS, SEA, or SEE for 24 h (TNF-α), 48 h (IL-1β), & 72 h (IFN-γ) | IL-1β & TNF-α concentrations in LPS-stimulated cells were enhanced by the addition of Zn in a concentration-dependent manner; Zn down-regulated levels of IL-1β & TNF-α in cells stimulated with SEA & SEE superantigens |
| Scuderi, 1990 [ 56] | PBMC were incubated with 30, 60, 120, 250, 500, 1000, 2000 µM Zn for 18 h; in addition, cells were incubated with 63, 125, 250, 500, 1000 µM Zn plus a substimulatory concentration of LPS (0.01 pg/mL) | Addition of Zn resulted in a concentration-dependent stimulation of TNF (with a peak at 250 µM) & IL-1β (peak at 120 µM), IL-6 was unaffected by Zn; Zn & LPS in combination resulted in a synergistic stimulation of TNF but not IL-1β secretion |
4. Immune Cells
5. Key Signaling Mechanisms: NF-κB and Nitric Oxide
6. Cardiometabolic Disease

7. Conclusions
Conflict of Interest
References
- Prasad, A.S.; Halsted, J.A.; Nadimi, M. Syndrome of iron deficiency anaemia, hepatosplenomegaly, hypogonadism, dwarfism and geophagia. Am. J. Med. 1961, 31, 532–546. [Google Scholar] [CrossRef]
- Vallee, B.L.; Falchuk, K.H. The biochemical basis of zinc physiology. Physiol. Rev. 1993, 73, 79–118. [Google Scholar] [CrossRef]
- Beyersmann, D.; Haase, H. Functions of zinc in signaling, proliferation and differentiation of mammalian cells. Biometals 2001, 14, 331–341. [Google Scholar] [CrossRef]
- Krężel, A.; Maret, W. Zinc-buffering capacity of a eukaryotic cell at physiological pZn. J. Biol. Inorg. Chem. 2006, 11, 1049–1062. [Google Scholar] [CrossRef]
- Frederickson, C.J.; Koh, J.Y.; Bush, A.I. The neurobiology of zinc in health and disease. Nat. Rev. Neurosci. 2005, 6, 449–462. [Google Scholar] [CrossRef]
- Ezzati, M.; Lopez, A.D.; Rodgers, A.; Vander Hoorn, S.; Murray, C.J.L. Comparative Risk Assessment Collaborating Group. Selected major risk factors and global and regional burden of disease. Lancet 2002, 360, 1347–1360. [Google Scholar]
- Wuehler, S.E.; Peerson, J.M.; Brown, K.H. Use of national food balance data to estimate the adequacy of zinc in national food supplies: Methodology and regional estimates. Public Health Nutr. 2005, 8, 812–819. [Google Scholar]
- Prasad, A.S. Zinc in human health: Effect of zinc on immune cells. Mol. Med. 2008, 14, 353–357. [Google Scholar] [CrossRef]
- Kruse-Jarres, J.D. The significance of zinc for humoral and cellular immunity. J. Trace Elem. Electrolytes Health Dis. 1989, 3, 1–8. [Google Scholar]
- Abbas, A.K.; Lichtman, A.H. Cellular and Molecular Immunology, 5th ed; Elsevier Saunders: Philadelphia, PA, USA, 2005. [Google Scholar]
- Zhou, X.; Fragala, M.S.; McElhaney, J.E.; Kuchel, G.A. Conceptual and methodological issues relevant to cytokine and inflammatory marker measurements in clinical research. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 541–547. [Google Scholar] [CrossRef]
- Medzhitov, R. Inflammation 2010: New adventures of an old flame. Cell 2010, 140, 771–776. [Google Scholar] [CrossRef]
- Nathan, C.; Ding, A. Nonresolving inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef]
- Krebs, N.E.; Hambidge, K.M. Zinc metabolism and homeostasis: The application of tracer techniques to human zinc physiology. Biometals 2001, 14, 397–412. [Google Scholar] [CrossRef]
- Foster, M.; Samman, S. Zinc and redox signaling: Perturbations associated with cardiovascular disease and diabetes mellitus. Antioxid. Redox Signal. 2010, 13, 1549–1573. [Google Scholar] [CrossRef]
- Prasad, A.S. Zinc and immunity. Mol. Cell. Biochem. 1998, 188, 63–69. [Google Scholar] [CrossRef]
- Besecker, B.Y.; Exline, M.C.; Hollyfield, J.; Phillips, G.; Disilvestro, R.A.; Wewers, M.D.; Knoell, D.L. A comparison of zinc metabolism, inflammation, and disease severity in critically ill infected and noninfected adults early after intensive care unit admission. Am. J. Clin. Nutr. 2011, 93, 1356–1364. [Google Scholar] [CrossRef]
- Egefjord, L.; Jensen, J.L.; Bang-Berthelsen, C.H.; Petersen, A.B.; Smidt, K.; Schmitz, O.; Karlsen, A.E.; Pociot, F.; Chimienti, F.; Rungby, J.; Magnusson, N.E. Zinc transporter gene expression is regulated by pro-inflammatory cytokines: A potential role for zinc transporters in beta-cell apoptosis? BMC Endocr. Disord. 2009, 9, 7. [Google Scholar] [CrossRef]
- Lang, C.; Murgia, C.; Leong, M.; Tan, L.W.; Perozzi, G.; Knight, D.; Ruffin, R.; Zalewski, P. Anti-inflammatory effects of zinc and alterations in zinc transporter mRNA in mouse models of allergic inflammation. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L577–L584. [Google Scholar]
- Liuzzi, J.P.; Lichten, L.A.; Rivera, S.; Blanchard, R.K.; Aydemir, T.B.; Knutson, M.D.; Ganz, T.; Cousins, R.J. Interleukin-6 regulates the zinc transporter Zip14 in liver and contributes to the hypozincemia of the acute-phase response. Proc. Natl. Acad. Sci. USA 2005, 102, 6843–6848. [Google Scholar]
- Cousins, R.J.; Leinart, A.S. Tissue-specific regulation of zinc metabolism and metallothionein genes by interleukin 1. FASEB J. 1988, 2, 2884–2890. [Google Scholar]
- De, S.K.; McMaster, M.T.; Andrews, G.K. Endotoxin induction of murine metallothionein gene expression. J. Biol. Chem. 1990, 265, 15267–15274. [Google Scholar]
- Lichten, L.A.; Cousins, R.J. Mammalian zinc transporters: Nutritional and physiological regulation. Annu. Rev. Nutr. 2009, 29, 153–176. [Google Scholar] [CrossRef]
- Kim, S.; Watanabe, K.; Shirahata, T.; Watarai, M. Zinc uptake system (znuA locus) of Brucella abortus is essential for intracellular survival and virulence in mice. J. Vet. Med. Sci. 2004, 66, 1059–1063. [Google Scholar] [CrossRef]
- Gaetke, L.M.; McClain, C.J.; Talwalkar, R.T.; Shedlofsky, S.I. Effects of endotoxin on zinc metabolism in human volunteers. Am. J. Physiol. 1997, 272, E952–E956. [Google Scholar]
- Young, B.; Ott, L.; Kasarskis, E.; Rapp, R.; Moles, K.; Dempsey, R.J.; Tibbs, P.A.; Kryscio, R.; McClain, C. Zinc supplementation is associated with improved neurologic recovery rate and visceral protein levels of patients with severe closed head injury. J. Neurotrauma 1996, 13, 25–34. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Costarelli, L.; Muti, E.; Malavolta, M.; Cipriano, C.; Giacconi, R.; Tesei, S.; Piacenza, F.; Pierpaoli, S.; Gasparini, N.; Faloia, E.; et al. Distinctive modulation of inflammatory and metabolic parameters in relation to zinc nutritional status in adult overweight/obese subjects. J. Nutr. Biochem. 2010, 21, 432–437. [Google Scholar] [CrossRef]
- Prasad, A.S.; Beck, F.W.; Grabowski, S.M.; Kaplan, J.; Mathog, R.H. Zinc deficiency: Changes in cytokine production and T-cell subpopulations in patients with head and neck cancer and in noncancer subjects. Proc. Assoc. Am. Physicians 1997, 109, 68–77. [Google Scholar]
- Beck, F.W.; Li, Y.; Bao, B.; Prasad, A.S.; Sarkar, F.H. Evidence for reprogramming global gene expression during zinc deficiency in the HUT-78 cell line. Nutrition 2006, 22, 1045–1056. [Google Scholar]
- Prasad, A.S.; Bao, B.; Beck, F.W.J.; Sarkar, F.H. Correction of interleukin-2 gene expression by in vitro zinc addition to mononuclear cells from zinc-deficient human subjects: A specific test for zinc deficiency in humans. Transl. Res. 2006, 148, 325–333. [Google Scholar] [CrossRef]
- Pinna, K.; Kelley, D.S.; Taylor, P.C.; King, J.C. Immune functions are maintained in healthy men with low zinc intake. J. Nutr. 2002, 132, 2033–2036. [Google Scholar]
- Wieringa, F.T.; Dijkhuizen, M.A.; West, C.E.; van der Ven-Jongekrijg, J.; van der Meer, J.W.; Muhilal. Reduced production of immunoregulatory cytokines in vitamin A- and zinc-deficient Indonesian infants. Eur. J. Clin. Nutr. 2004, 58, 1498–1504. [Google Scholar] [CrossRef]
- Bao, B.; Prasad, A.S.; Beck, F.W.J.; Snell, D.; Suneja, A.; Sarkar, F.H.; Doshi, N.; Fitzgerald, J.T.; Swerdlow, P. Zinc supplementation decreases oxidative stress, incidence of infection, and generation of inflammatory cytokines in sickle cell disease patients. Transl. Res. 2008, 152, 67–80. [Google Scholar] [CrossRef]
- Prasad, A.S.; Beck, F.W.J.; Bao, B.; Fitzgerald, J.T.; Snell, D.C.; Steinberg, J.D.; Cardozo, L.J. Zinc supplementation decreases incidence of infections in the elderly: Effect of zinc on generation of cytokines and oxidative stress. Am. J. Clin. Nutr. 2007, 85, 837–844. [Google Scholar]
- Bao, B.; Prasad, A.S.; Beck, F.W.J.; Fitzgerald, J.T.; Snell, D.; Bao, G.W.; Singh, T.; Cardozo, L.J. Zinc decreases C-reactive protein, lipid peroxidation, and inflammatory cytokines in elderly subjects: A potential implication of zinc as an atheroprotective agent. Am. J. Clin. Nutr. 2010, 91, 1634–1641. [Google Scholar] [CrossRef]
- Prasad, A.S.; Bao, B.; Beck, F.W.J.; Kucuk, O.; Sarkar, F.H. Antioxidant effect of zinc in humans. Free Radic. Biol. Med. 2004, 37, 1182–1190. [Google Scholar] [CrossRef]
- Raqib, R.; Roy, S.K.; Rahman, M.J.; Azim, T.; Ameer, S.S.; Chisti, J.; Andersson, J. Effect of zinc supplementation on immune and inflammatory responses in pediatric patients with shigellosis. Am. J. Clin. Nutr. 2004, 79, 444–450. [Google Scholar]
- Sandstead, H.H.; Prasad, A.S.; Penland, J.G.; Beck, F.W.J.; Kaplan, J.; Egger, N.G.; Alcock, N.W.; Carroll, R.M.; Ramanujam, V.M.S.; Dayal, H.H.; et al. Zinc deficiency in Mexican American children: Influence of zinc and other micronutrients on T cells, cytokines, and antiinflammatory plasma proteins. Am. J. Clin. Nutr. 2008, 88, 1067–1073. [Google Scholar]
- Aydemir, T.B.; Blanchard, R.K.; Cousins, R.J. Zinc supplementation of young men alters metallothionein, zinc transporter, and cytokine gene expression in leukocyte populations. Proc. Natl. Acad. Sci. USA 2006, 103, 1699–1704. [Google Scholar]
- Kahmann, L.; Uciechowski, P.; Warmuth, S.; Plümäkers, B.; Gressner, A.M.; Malavolta, M.; Mocchegiani, E.; Rink, L. Zinc supplementation in the elderly reduces spontaneous inflammatory cytokine release and restores T cell functions. Rejuvenation Res. 2008, 11, 227–237. [Google Scholar] [CrossRef]
- Mariani, E.; Cattini, L.; Neri, S.; Malavolta, M.; Mocchegiani, E.; Ravaglia, G.; Facchini, A. Simultaneous evaluation of circulating chemokine and cytokine profiles in elderly subjects by multiplex technology: Relationship with zinc status. Biogerontology 2008, 7, 449–459. [Google Scholar]
- Mocchegiani, E.; Giacconi, R.; Costarelli, L.; Muti, E.; Cipriano, C.; Tesei, S.; Pierpaoli, S.; Giuli, C.; Papa, R.; Marcellini, F.; et al. Zinc deficiency and IL-6–174G/C polymorphism in old people from different European countries: Effect of zinc supplementation. ZINCAGE study. Exp. Gerontol. 2008, 43, 433–444. [Google Scholar] [CrossRef]
- Kara, E.; Ozal, M.; Gunay, M.; Kilic, M.; Baltaci, A.K.; Mogulkoc, R. Effects of exercise and zinc supplementation on cytokine release in young wrestlers. Biol. Trace Elem. Res. 2011, 143, 1435–1440. [Google Scholar] [CrossRef]
- Bao, B.; Prasad, A.S.; Beck, F.W.; Bao, G.W.; Singh, T.; Ali, S.; Sarkar, F.H. Intracellular free zinc up-regulates IFN-γ and T-bet essential for Th1 differentiation in Con-A stimulated HUT-78 cells. Biochem. Biophys. Res. Commun. 2011, 407, 703–707. [Google Scholar] [CrossRef]
- Hayashi, K.; Ishizuka, S.; Yokoyama, C.; Hatae, T. Attenuation of interferon-gamma mRNA expression in activated Jurkat T cells by exogenous zinc via down-regulation of the calcium-independent PKC-AP-1 signaling pathway. Life Sci. 2008, 83, 6–11. [Google Scholar] [CrossRef]
- Prasad, A.S.; Bao, B.; Beck, F.W.J.; Sarkar, F.H. Zinc-suppressed inflammatory cytokines by induction of A20-mediated inhibition of nuclear factor-κB. Nutrition 2011, 27, 816–823. [Google Scholar] [CrossRef]
- Freitas, M.; Fernandes, E. Zinc, cadmium and nickel increase the activation of NF-κB and the release of cytokines from THP-1 monocytic cells. Metallomics 2011, 3, 1238–1243. [Google Scholar] [CrossRef]
- Tsou, T.C.; Chao, H.R.; Yeh, S.C.; Tsai, F.Y.; Lin, H.J. Zinc induces chemokine and inflammatory cytokine release from human promonocytes. J. Hazard. Mater. 2011, 196, 335–341. [Google Scholar]
- Metz, C.H.; Schröder, A.K.; Overbeck, S.; Kahmann, L.; Plümäkers, B.; Rink, L. T-helper type 1 cytokine release is enhanced by in vitro zinc supplementation due to increased natural killer cells. Nutrition 2007, 23, 157–163. [Google Scholar] [CrossRef]
- Poleganov, M.A.; Pfeilschifter, J.; Mühl, H. Expanding extracellular zinc beyond levels reflecting the albumin-bound plasma zinc pool potentiates the capability of IL-1β, IL-18, and IL-12 to act as IFN-γ-inducing factors on PBMC. J. Interferon Cytokine Res. 2007, 27, 997–1001. [Google Scholar] [CrossRef]
- Chang, K.L.; Hung, T.C.; Hsieh, B.S.; Chen, Y.H.; Chen, T.F.; Cheng, H.L. Zinc at pharmacologic concentrations affects cytokine expression and induces apoptosis of human peripheral blood mononuclear cells. Nutrition 2006, 22, 465–474. [Google Scholar] [CrossRef]
- Von Bülow, V.; Rink, L.; Haase, H. Zinc-mediated inhibition of cyclic nucleotide phosphodiesterase activity and expression suppresses TNF-alpha and IL-1 beta production in monocytes by elevation of guanosine 3′,5′-cyclic monophosphate. J. Immunol. 2005, 175, 4697–4705. [Google Scholar]
- Wellinghausen, N.; Martin, M.; Rink, L. Zinc inhibits interleukin-1-dependent T cell stimulation. Eur. J. Immunol. 1997, 27, 2529–2535. [Google Scholar] [CrossRef]
- Driessen, C.; Hirv, K.; Wellinghausen, N.; Kirchner, H.; Rink, L. Influence of serum on zinc, toxic shock syndrome toxin-1, and lipopolysaccharide-induced production of IFN-gamma and IL-1 beta by human mononuclear cells. J. Leukoc. Biol. 1995, 57, 904–908. [Google Scholar]
- Scuderi, P. Differential effects of copper and zinc on human peripheral blood monocyte cytokine secretion. Cell. Immunol. 1990, 126, 391–405. [Google Scholar] [CrossRef]
- Prasad, A.S. Zinc: Mechanisms of host defense. J. Nutr. 2007, 137, 1345–1349. [Google Scholar]
- Beck, F.W.J.; Prasad, A.S.; Kaplan, J.; Fitzgerald, J.T.; Brewer, G.J. Changes in cytokine production and T cell subpopulations in experimentally induced zinc-deficient humans. Am. J. Physiol. 1997, 272, E1002–E1007. [Google Scholar]
- Tapazoglou, E.; Prasad, A.S.; Hill, G.; Brewer, G.J.; Kaplan, J. Decreased natural killer cell activity in zinc deficient subjects with sickle cell disease. J. Lab. Clin. Med. 1985, 105, 19–22. [Google Scholar]
- Fantone, J.C.; Ward, P.A. Polymorphonuclear leukocyte-mediated cell and tissue injury: Oxygen metabolites and their relations to human disease. Hum. Pathol. 1985, 16, 973–978. [Google Scholar] [CrossRef]
- Scapini, P.; Lapinet-Vera, J.A.; Gasperini, S.; Calzetti, F.; Bazzoni, F.; Cassatella, M.A. The neutrophil as a cellular source of chemokines. Immunol. Rev. 2000, 177, 195–203. [Google Scholar]
- Finamore, A.; Massimi, M.; Conti Devirgiliis, L.; Mengheri, E. Zinc deficiency induces membrane barrier damage and increases neutrophil transmigration in Caco-2 cells. J. Nutr. 2008, 138, 1664–1670. [Google Scholar]
- Gavella, M.; Lipovac, V.; Car, A. In vitro effect of zinc on superoxide anion production by polymorphonuclear leukocytes of diabetic patients. Acta Diabetol. 2000, 37, 135–137. [Google Scholar] [CrossRef]
- Siminiak, T.; Flores, N.A.; Sheridan, D.J. Neutrophil interactions with endothelium and platelets: Possible role in the development of cardiovascular injury. Eur. Heart J. 1995, 16, 160–170. [Google Scholar]
- Verstrepen, L.; Bekaert, T.; Chau, T.L.; Tavernier, J.; Chariot, A.; Beyaert, R. TLR-4, IL-1R and TNF-R signaling to NF-κB: Variations on a common theme. Cell. Mol. Life Sci. 2008, 65, 2964–2978. [Google Scholar] [CrossRef]
- Bao, B.; Prasad, A.; Beck, F.W.; Suneja, A.; Sarkar, F. Toxic effect of zinc on NF-kappaB, IL-2, IL-2 receptor alpha, and TNF-alpha in HUT-78 (Th(0)) cells. Toxicol. Lett. 2006, 166, 222–228. [Google Scholar] [CrossRef]
- von Bülow, V.; Dubben, S.; Engelhardt, G.; Hebel, S.; Plümäkers, B.; Heine, H.; Rink, L.; Haase, H. Zinc-dependent suppression of TNF-α production is mediated by protein kinase A-induced inhibition of Raf-1, IκB Kinase β, and NF-κB. J. Immunol. 2007, 179, 4180–4186. [Google Scholar]
- Song, H.Y.; Rothe, M.; Goeddel, D.V. The tumor necrosis factor-inducible zinc finger protein A20 interacts with TRAF1/TRAF2 and inhibits NF-κB activation. Proc. Natl. Acad. Sci. USA 1996, 93, 6721–6725. [Google Scholar]
- Kim, Y.M.; Cao, D.; Reed, W.; Wu, W.; Jaspers, I.; Tal, T.; Bromberg, P.A.; Samet, J.M. Zn2+-induced NF-κB-dependent transcriptional activity involves site-specific p65/RelA phosphorylation. Cell. Signal. 2007, 19, 538–546. [Google Scholar] [CrossRef]
- Bao, S.; Liu, M.J.; Lee, B.; Besecker, B.; Lai, J.P.; Guttridge, D.C.; Knoell, D.L. Zinc modulates the innate immune response in vivo to polymicrobial sepsis through regulation of NF-κB. Am. J. Physiol. Lung Cell. Mol. Physiol. 2010, 298, L744–L754. [Google Scholar] [CrossRef]
- Dinarello, C.A. Immunological and inflammatory functions of the Interleukin-1 family. Annu. Rev. Immunol. 2009, 27, 519–550. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef]
- Zou, M.H.; Shi, C.; Cohen, R.A. Oxidation of the zinc-thiolate complex and uncoupling of endothelial nitric oxide synthase by peroxynitrite. J. Clin. Invest. 2002, 109, 817–826. [Google Scholar]
- Thomas, S.R.; Witting, P.K.; Drummond, G.R. Redox control of endothelial function and dysfunction: Molecular mechanisms and therapeutic opportunities. Antioxid. Redox Signal. 2008, 10, 1713–1765. [Google Scholar] [CrossRef]
- Xu, J.; Xie, Z.; Reece, R.; Pimental, D.; Zou, M.H. Uncoupling of endothelial nitric oxidase synthase by hypochlorous acid. Role of NAD(P)H oxidase-derived superoxide and peroxynitrite. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 2688–2695. [Google Scholar] [CrossRef]
- Azumi, H.; Inoue, N.; Takeshita, S.; Rikitake, Y.; Kawashima, S.; Hayashi, Y.; Itoh, H.; Yokoyama, M. Expression of NADH/NADPH oxidase p22phox in human coronary arteries. Circulation 1999, 100, 1494–1498. [Google Scholar] [CrossRef]
- Matsunaga, Y.; Kawai, Y.; Kohda, Y.; Gemba, M. Involvement of activation of NADPH oxidase and extracellular signal-regulated kinase (ERK) in renal cell injury induced by zinc. J. Toxicol. Sci. 2005, 30, 135–144. [Google Scholar] [CrossRef]
- Foster, M.; Samman, S. Zinc and Atherosclerosis: Clinical Observations and Potential Mechanisms. In Zinc and Human Health: Biomedical and Health Research; Rink, L., Ed.; IOS Press: Amsterdam, The Netherlands, 2011; Volume 76, pp. 347–372. [Google Scholar]
- Rowe, D.J.; Bobilya, D.J. Albumin facilitates zinc acquisition by endothelial cells. Proc. Soc. Exp. Biol. Med. 2000, 224, 178–186. [Google Scholar] [CrossRef]
- Beattie, J.H.; Kwun, I.S. Is zinc deficiency a risk factor for atherosclerosis? Br. J. Nutr. 2004, 91, 177–181. [Google Scholar] [CrossRef]
- Hughes, S.; Samman, S. The effect of zinc supplementation in humans on plasma lipids, antioxidant status, and thrombogenesis. J. Am. Coll. Nutr. 2006, 25, 285–291. [Google Scholar]
- Foster, M.; Petocz, P.; Samman, S. Effects of zinc on plasma lipoprotein cholesterol concentrations in humans: A meta-analysis of randomised controlled trials. Atherosclerosis 2010, 210, 344–352. [Google Scholar] [CrossRef]
- Meerarani, P.; Ramadass, P.; Toborek, M.; Bauer, H.C.; Bauer, H.; Hennig, B. Zinc protects against apoptosis of endothelial cells induced by linoleic acid and tumor necrosis factor alpha. Am. J. Clin. Nutr. 2000, 71, 81–87. [Google Scholar]
- Wiseman, D.A.; Wells, S.M.; Hubbard, M.; Welker, J.E.; Black, S.M. Alterations in zinc homeostasis underlie endothelial cell death induced by oxidative stress from acute exposure to hydrogen peroxide. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L165–L177. [Google Scholar]
- Hennig, B.; Meerarani, P.; Toborek, M.; McClain, C.J. Antioxidant-like properties of zinc in activated endothelial cells. J. Am. Coll. Nutr. 1999, 18, 152–158. [Google Scholar]
- Bourcier, T.; Sukhova, G.; Libby, P. The nuclear factor kB signaling pathway participates in dysregulation of vascular smooth muscle cells in vitro and in human atherosclerosis. J. Biol. Chem. 1997, 272, 15817–15824. [Google Scholar]
- De Martin, R.; Hoeth, M.; Hofer-Warbinek, R.; Schmid, J.A. The transcription factor NF-κB and the regulation of vascular cell function. Arterioscler. Thromb. Vasc. Biol. 2000, 20, e83–e88. [Google Scholar] [CrossRef]
- Pasterkamp, G.; van Keulen, J.K.; de Kleijn, D.P.V. Role of Toll-like receptor 4 in the initiation and progression of atherosclerotic disease. Eur. J. Clin. Invest. 2004, 34, 328–334. [Google Scholar] [CrossRef]
- Cortese-Krott, M.M.; Suschek, C.V.; Wetzel, W.; Kröncke, K.D.; Kolb-Bachofen, V. Nitric oxide-mediated protection of endothelial cells from hydrogen peroxide is mediated by intracellular zinc and glutathione. Am. J. Physiol. Cell Physiol. 2009, 296, 811–820. [Google Scholar] [CrossRef]
- Donath, M.Y.; Størling, J.; Maedler, K.; Mandrup-Poulsen, T. Inflammatory mediators and islet beta-cell failure: A link between type 1 and type 2 diabetes. J. Mol. Med. 2003, 81, 455–470. [Google Scholar] [CrossRef]
- Maedler, K.; Sergeev, P.; Ris, F.; Oberholzer, J.; Joller-Jemelka, H.I.; Spinas, G.A.; Kaiser, N.; Halban, P.A.; Donath, M.Y. Glucose-induced beta cell production of IL1-beta contributes to glucotoxicity in human pancreatic islets. J. Clin. Invest. 2002, 110, 851–860. [Google Scholar]
- Ehses, J.A.; Perren, A.; Eppler, E.; Ribaux, P.; Pospisilik, J.A.; Maor-Cahn, R.; Gueripel, X.; Ellingsgaard, H.; Schneider, M.K.; Biollaz, G.; et al. Increased number of islet-associated macrophages in type 2 diabetes. Diabetes 2007, 56, 2356–2370. [Google Scholar]
- Spranger, J.; Kroke, A.; Möhlig, M.; Hoffman, K.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F. Inflammatory cytokines and the risk to develop type 2 diabetes: Results of the prospective population-based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Diabetes 2003, 52, 812–817. [Google Scholar] [CrossRef]
- Donath, M.Y.; Böni-Schnetzler, M.; Ellingsgaard, H.; Halban, P.A.; Ehses, J.A. Cytokine production by islets in health and diabetes: Cellular origin, regulation and function. Trends Endocrinol. Metab. 2010, 21, 261–267. [Google Scholar] [CrossRef]
- Arai, K.; Iizuka, S.; Tada, Y.; Oikawa, K.; Taniguchi, N. Increase in the glucosylated form of erythrocyte Cu-Zn-superoxide dismutase in diabetes and close association of the nonenzymatic glucosylation with the enzyme activity. Biochim. Biophys. Acta 1987, 924, 292–296. [Google Scholar] [CrossRef]
- King, G.L.; Loeken, M.R. Hyperglycemia-induced oxidative stress in diabetic complications. Histochem. Cell Biol. 2004, 122, 333–338. [Google Scholar] [CrossRef]
- Gupta, R.; Garg, V.K.; Mathur, D.K.; Goyal, R.K. Oral zinc therapy in diabetic neuropathy. J. Assoc. Physicians India 1998, 46, 939–942. [Google Scholar]
- Kajanachumpol, S.; Srisurapanon, S.; Supanit, I.; Roongpisuthipong, C.; Apibal, S. Effect of zinc supplementation on zinc status, copper status and cellular immunity in elderly patients with diabetes mellitus. J. Med. Assoc. Thail. 1995, 78, 344–349. [Google Scholar]
- Samman, S.; Foster, M.; Capdor, J.; Stathakis, E. Zinc, diabetes mellitus and cardiovascular disease in humans: A critical appraisal. In Proceedings of International Society for Zinc Biology Conference, Melbourne, Australia, 15–19 January 2012. Abstract #71.
- Wellinghausen, N.; Kirchner, H.; Rink, L. The immunobiology of zinc. Immunol. Today 1997, 18, 519–521. [Google Scholar] [CrossRef]
- Cave, A.C.; Brewer, A.C.; Narayanapanicker, A.; Ray, R.; Grieve, D.J.; Walker, S.; Shah, A.M. NADPH oxidases in cardiovascular health and disease. Antioxid. Redox Signal. 2006, 8, 691–728. [Google Scholar] [CrossRef] [Green Version]
- Ibs, K.H.; Rink, L. Zinc-altered immune function. J. Nutr. 2003, 133, 1452–1456. [Google Scholar]
- Samman, S. Dietary versus cellular zinc: The antioxidant paradox. Free Radic. Biol. Med. 1993, 14, 95–96. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Foster, M.; Samman, S. Zinc and Regulation of Inflammatory Cytokines: Implications for Cardiometabolic Disease. Nutrients 2012, 4, 676-694. https://doi.org/10.3390/nu4070676
Foster M, Samman S. Zinc and Regulation of Inflammatory Cytokines: Implications for Cardiometabolic Disease. Nutrients. 2012; 4(7):676-694. https://doi.org/10.3390/nu4070676
Chicago/Turabian StyleFoster, Meika, and Samir Samman. 2012. "Zinc and Regulation of Inflammatory Cytokines: Implications for Cardiometabolic Disease" Nutrients 4, no. 7: 676-694. https://doi.org/10.3390/nu4070676