Article Text

Abstract
Objectives Homocysteine has been causally associated with various adverse health outcomes. Evidence supporting the relationship between lead and homocysteine levels has been accumulating, but most prior studies have not focused on the interaction with genetic polymorphisms.
Methods From a community-based prospective cohort, we analysed 386 participants (aged 41–71 years) with information regarding blood lead and plasma homocysteine levels. Blood lead levels were measured between 2001 and 2003, and plasma homocysteine levels were measured in 2007. Interactions of lead levels with 42 genotyped single-nucleotide polymorphisms (SNPs) in five genes (TF, HFE, CBS, BHMT and MTR) were assessed via a 2-degree of freedom (df) joint test and a 1-df interaction test. In secondary analyses using imputation, we further assessed 58 imputed SNPs in the TF and MTHFR genes.
Results Blood lead concentrations were positively associated with plasma homocysteine levels (p=0.0276). Six SNPs in the TF and MTR genes were screened using the 2-df joint test, and among them, three SNPs in the TF gene showed interactions with lead with respect to homocysteine levels through the 1-df interaction test (p<0.0083). Seven SNPs in the MTHFR gene were associated with homocysteine levels at an α-level of 0.05, but the associations did not persist after Bonferroni correction. These SNPs did not show interactions with lead levels.
Conclusions Blood lead levels were positively associated with plasma homocysteine levels measured 4-6 years later, and three SNPs in the TF gene modified the association.
- Blood Lead Levels
- Homocysteine
- Gene-environment Interaction
- Single-nucleotide Polymorphism
Statistics from Altmetric.com
What this paper adds
Homocysteine has been causally associated with various adverse health outcomes, such as Alzheimer’s disease and schizophrenia.
Although accumulating evidence supports the association between lead and homocysteine, most previous studies did not focus on interactions between lead and genetic polymorphisms, with respect to homocysteine levels.
Blood lead levels were positively associated with plasma homocysteine levels measured 4–6 years later, and single-nucleotide polymorphisms (SNPs) in the TF gene modified the association between lead and homocysteine levels in a community-based prospective cohort.
These results suggest that iron distribution may influence the effect of lead toxicity on homocysteine levels by affecting homocysteine metabolism.
The present data show the importance of continued efforts to reduce human lead exposure and suggest a direction for future mechanistic studies.
Introduction
Homocysteine, a sulfur-containing amino acid generated from methionine, has been identified as a causal factor in various adverse health outcomes. Maternal homocysteine levels during pregnancy were inversely associated with birth weights in Mendelian randomisation analyses.1 2 In terms of neurological outcomes, increased homocysteine levels were found to be causally related to Alzheimer’s disease3 and schizophrenia4 in previous studies. Prior findings have also demonstrated that homocysteine is an independent risk factor for cardiovascular and metabolic diseases,5–7 whereas causal associations with type 2 diabetes or blood pressure are not clear.8–10
Alcohol consumption, tobacco smoking,11 inadequate diet and micronutrient levels,12 13 ageing and decreased renal function14 15 have been indicated as predictors of plasma homocysteine levels. In recent years, some studies have reported an association between lead exposure and increased homocysteine levels.16–21 Previous studies provided evidence suggesting that even small increases in lead levels are associated with an increase in homocysteine levels sufficient to cause extensive changes in the prevalence of related adverse health outcomes and diseases.16 20
Several enzymes in the homocysteine metabolism pathway, such as cystathionine beta-synthase (CBS), have been suggested to play important roles in the biological pathways between lead exposure and homocysteine levels.17 19 20 Investigation of the biological mechanisms is important for establishing a potential causal relationship between lead exposure and homocysteine levels, and gene–environment interaction studies are effective tools for assessing hypothesised biological mechanisms and advancing the mechanistic understanding.22 However, despite their importance, studies investigating the biological pathways of lead–homocysteine associations, including gene–environment interactions, have been rare.
In addition, genetic susceptibility to environmental exposure for health outcomes of interest can be evaluated by assessing gene–environment interactions rather than assessing the main effects of genetic polymorphisms on exposure or outcome levels.22–24 Therefore, in the present study, we investigated interactions between blood lead levels, an indicator of lead exposure, and targeted genetic polymorphisms related to homocysteine metabolism with respect to homocysteine levels.
Methods
Study population
The present study was performed using data from the ongoing population-based cohort in the Ansung and Ansan areas (Republic of Korea), established as part of the Korean Genome and Epidemiology Study (KoGES). Details of the Ansung–Ansan cohort have been described elsewhere.25 Briefly, in the Ansung–Ansan cohort, 10 038 adults were recruited between June 2001 and January 2003, using a two-stage cluster sampling method and followed up with every 2 years. At the time of each survey, a comprehensive health examination, blood sampling and interview using a structured questionnaire were conducted. Genotype data were obtained for 8842 participants from 10 004 available samples after completion of a quality control process.26
Blood lead concentrations were analysed for 575 randomly selected blood samples, which were collected between June 2001 and October 2001 (at the baseline survey; n=499) or April 2003 (at the second survey; n=76). Among 575 participants with blood lead information, plasma homocysteine levels were available for 386 participants at the time of the fourth survey (March 2007–December 2007).
Ethics, consent and permissions
The Institutional Review Board of Seoul National University Hospital approved the study protocol (C-1306-046-495), and all study participants provided written informed consent to participate.
Genotyping and SNP selection
Peripheral blood was sampled for genomic DNA isolation. Genotypes for the target gene single-nucleotide polymorphisms (SNPs) were obtained using the Affymetrix Genome-Wide Human Array 5.0 (Affymetrix, Santa Clara, California, USA) with standard quality control procedures to assess the Hardy-Weinberg equilibria (p<10−6), genotype call rates (<95%) and minor allele frequencies (<0.01), as previously described.26 Imputation of untyped or missing SNPs was conducted using IMPUTE,27 with International HapMap Phase II HCB and JPT genotype information used as a reference. The imputation methods and procedures were described in detail elsewhere.28
Six genes encoding CBS, transferrin (TF), haemochromatosis protein (HFE), betaine-homocysteine S-methyltransferase (BHMT), methionine synthase (MTR) and methylenetetrahydrofolate reductase (MTHFR), which are members of homocysteine metabolism pathways or were targeted as potential mediators between lead and homocysteine levels in previous studies,16 19 20 29–31 were selected and assessed in the present study. SNPs in these six genes were identified by searching the National Center for Biotechnology Information (NCBI) database. Among them, 42 SNPs in five genes (15 SNPs in TF, two in HFE, three in CBS, two in BHMT and 20 in MTR) available on the Affymetrix chip were analysed in this study (see table S1 in the online supplementary file 1). Our goal was to investigate gene–environment interactions and not the effects of genetic variants on homocysteine levels. Therefore, we did not choose SNPs reported to be associated with homocysteine levels but evaluated available SNPs in the target genes that potentially interact with blood lead levels. We also assessed 30 imputed SNPs in the TF gene and 28 imputed SNPs in the MTHFR gene in our secondary analysis (see table S1 in the online supplementary file 1).
Supplementary file 1
Lead and homocysteine analyses
Venous blood was obtained from each participant after over 8 hours of fasting and collected in an EDTA tube. Blood lead concentrations were determined using graphite furnace atomic absorption spectrometry with Zeeman background correction (SpectrAA-800, Varian, Belrose, NSW, Australia).
To analyse homocysteine levels, samples were centrifuged for 5 min, and the plasma was placed on ice and kept in a refrigerator until analysis. The total plasma homocysteine concentration was determined using the ADVIA Centaur Immunoassay (Bayer, Leverkusen, Germany).
Statistical analysis
Homocysteine levels were log-transformed to approximate the normal distribution. Lead levels categorised into quartiles were used in the main and interaction analyses. We assessed the association of lead levels measured between 2001 and 2003 with homocysteine levels measured in 2007, using linear regression models adjusted for potential confounders selected a priori based on previous studies and an assumed biological pathway.16 18–21 The potential confounders included the age at the time of lead measurement (years), sex, alcohol intake (non-drinker, ex-drinker or current drinker), tobacco smoking (non-smoker, ex-smoker or current smoker), education level (less than high school, completed high school or more than high school), body mass index (kg/m2), time from lead measurement to homocysteine measurement (year) and creatinine clearance (mL/min). We estimated creatinine clearance based on the age (year), weight (kg) and serum creatinine level (mg/dL), using the equation: ((140−age) × weight)/(72×serum creatinine concentration] (×0.85, if the participant was female).16 32 In the sensitivity analysis, we also conducted analyses further adjusted for the levels of dietary vitamin B6, folate and both.
We evaluated interactions between lead levels and genetic polymorphisms with respect to homocysteine levels in two steps. First, we conducted a 2-degree of freedom (df) joint test, which is a robust screening method for gene discovery,23 33 for 42 targeted SNPs in five genes (TF, HFE, CBS, BHMT and MTR). The 2-df joint test was used to simultaneously evaluate the main genetic effects and gene–environment interactions. To test 2-df statistics for each SNP, we performed likelihood ratio tests between the full linear regression model containing covariates, SNP, lead levels multiplicative interaction term for each SNP and lead concentrations and the reduced model excluding SNP and the interaction term. Second, for the SNPs screened via 2-df joint tests at a significance level of 0.05, we further conducted conventional 1-df interaction tests to evaluate the gene–environment interactions per se,23 by testing the multiplicative interaction terms added to the adjusted linear regression models. In the second step, we applied the Bonferroni correction to consider the inflated α error and set the level of significance at p=0.0083. The additive genetic model, which has been demonstrated to show good performance even in cases where the true genetic model is unknown and the model specification is incorrect,34 35 was used in this analysis. However, we presented the results of stratified analyses by genotype based on the presence or absence of the variant allele (homozygous wild-type allele vs. heterozygote and homozygous variants combined), because only a small proportion of study participants was homozygous for the variant allele, leading to an unstable statistical estimation in this stratum. We also conducted non-parametric analysis using generalised additive models separated by the presence or absence of a variant allele of these SNPs to consider potential non-linear associations between lead and homocysteine levels in each stratum. For the secondary analysis, we performed the same procedure with 30 imputed SNPs in the TF gene and 28 in the MTHFR gene.
We used SAS V.9.4 (SAS, Cary, North Carolina, USA), R V.3.2.5 (The Comprehensive R Archive Network, Vienna, Austria: http://cran.r-project.org), PLINK V.1.07 (http://pngu.mgh.harvard.edu/~purcell/plink/) and Haploview V.4.2 (http://www.broadinstitute.org/haploview/) in the present analyses.
Results
Of the 386 participants with lead and homocysteine information, the age when the lead levels were measured ranged between 41 and 71 years (mean, 55 years). Among the participants, 205 (53.1%) were women, 229 (60.3%) had an education level of less than a high school degree, 160 (42.3%) were current drinker and 97 (25.7%) were current smokers. With respect to disease, 61 (15.8%) were diagnosed with hypertension, 35 (9.1%) with diabetes mellitus, 14 (3.6%) with thyroid disease and 9 (2.3%) experienced one or more myocardial infarction. Of the 378 participants with information regarding occupation, 126 (33.3%) were farmers, 113 (29.9%) were housewives, 38 (10.1%) were self-employed and 23 (6.1%) were factory workers. Lifestyle factors such as alcohol drinking and tobacco smoking did not change substantially during the study period (2001–2007). None of the participants were biologically related.
The mean (SD) blood lead concentration was 4.4 (1.9) µg/dL and the geometric mean (geometric SD) of the plasma homocysteine concentrations was 11.1 (1.4) µmol/L. Lead levels were higher in men, alcohol drinkers and tobacco smokers, and homocysteine levels were higher in the elderly, men, alcohol drinkers and tobacco smokers (table 1).
General characteristics of study participants (n=386)
Blood lead concentrations were positively associated with plasma homocysteine levels (table 2). When we further adjusted the models for the levels of dietary vitamin B6, folate and both, the results were similar, but had wider CI (see table S2 in the online supplementary file 1).
Associations between blood lead concentrations and plasma homocysteine levels (µmol/L)*
The 2-df joint tests were conducted for 42 SNPs in five targeted genes. Among them, four SNPs in the TF gene and two SNPs in the MTR gene were screened at a significance level of 0.05. When we conducted 1-df interaction tests for the six SNPs screened in the previous step, three SNPs in the TF gene (rs2715632, rs2715631 and rs2715627) showed interactions with lead levels after Bonferroni correction (p<0.0083) (table 3; table S3 in the online supplementary file 1). Stratified analyses by the number of variant alleles (0 vs 1 or 2) of these SNPs showed that positive associations between lead and homocysteine levels were only found among participants with variant alleles (table S4 in the online supplementary file 1; figure 1).

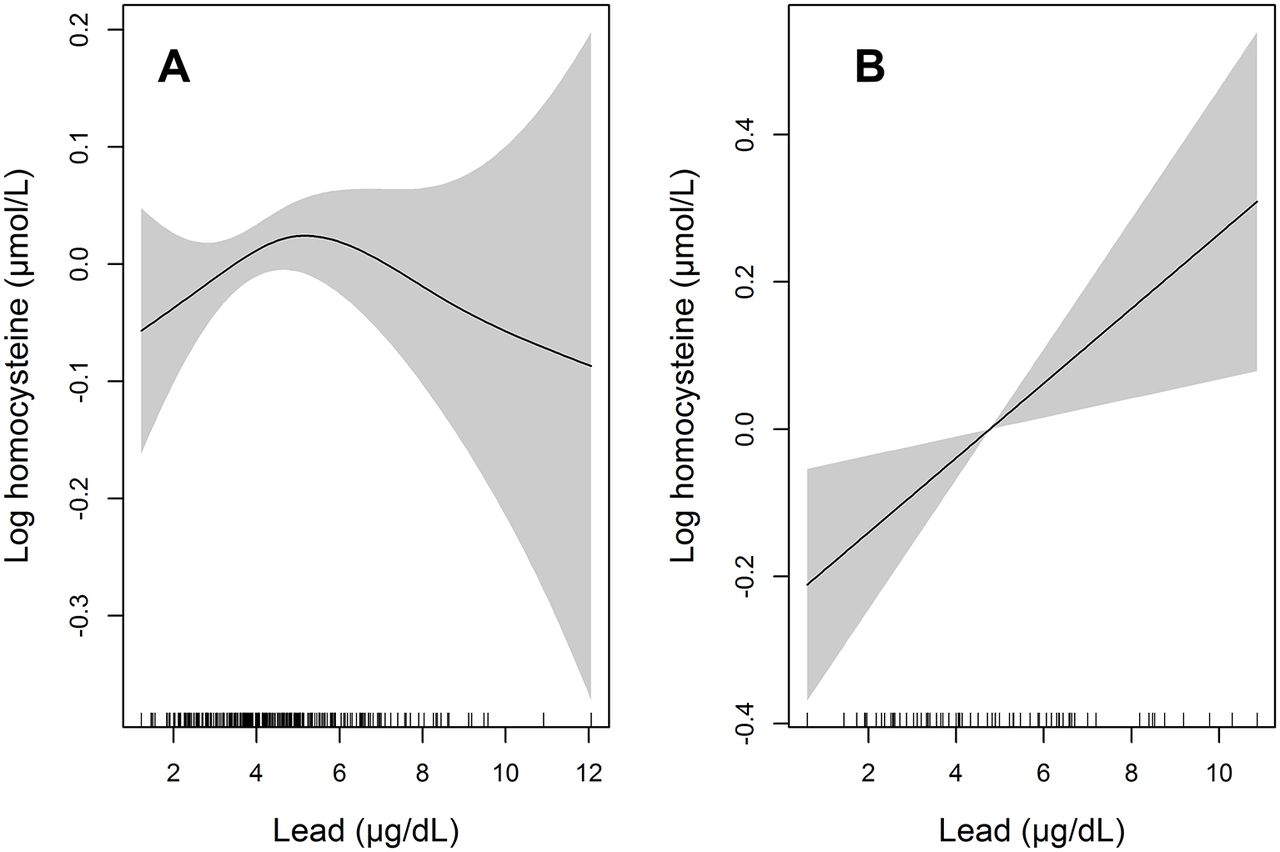{kind=link}
Penalised regression spline of blood lead levels on log-transformed homocysteine levels in participants without the rs2715627 variant C allele (A) and with the variant C allele (B). Solid lines represent spline curves and shaded areas represent 95% CIs. The models were adjusted for age at the time of lead measurement, sex, alcohol drinking, tobacco smoking, education level, body mass index, time from lead measurement to homocysteine measurement and creatinine clearance.
Single-nucleotide polymorphisms (SNPs) found to interact with lead levels versus homocysteine levels by the 2-df joint test
When we performed secondary analyses with 30 imputed SNPs in the TF gene and 28 imputed SNPs in the MTHFR gene, four SNPs in the TF gene and seven SNPs in the MTHFR gene were screened by the 2-df joint test. However, none of these SNPs showed evidence of interactions with lead levels by the 1-df interaction test after Bonferroni correction, while p Values for interaction of the four TF gene SNPs were lower than the 0.05 significance level (p=0.0084). These four imputed SNPs and selected three genotyped SNPs in the TF gene are in near physical distance and in the same linkage disequilibrium block (see table S1 in the online supplementary file 1). In contrast, the seven SNPs in the MTHFR gene were associated with homocysteine levels at an α-level of 0.05, but the associations did not persist after Bonferroni correction. These SNPs in the MTHFR gene did not show interactions with lead levels, with respect to homocysteine levels (see table S5 in the online supplementary file 1).
Discussion
We found positive associations between blood lead levels and plasma homocysteine levels measured 4–6 years later, using a community-based prospective cohort. Three SNPs (rs2715632, rs2715631 and rs2715627) in the TF gene interacted with lead levels with respect to homocysteine levels.
The lead and homocysteine levels measured in the present study were similar to or slightly higher than those found in previous studies conducted in elderly populations in the USA,16 20 but were much lower than those found in studies conducted in Asian populations residing in contaminated cities.18 21 In the current study, homocysteine levels were estimated to increase by 4.6% for every 3 µg/dL increase in blood lead levels, which is consistent with the results of a previous study conducted in elderly individuals in the USA16 This change in homocysteine levels associated with lead exposure can have public health implications considering the causal role of homocysteine in various health outcomes.1–4 The present results also demonstrate the importance of reducing lead exposure even in developed countries where lead levels are already low.
Blood lead levels were used as an indicator of lead exposure in the present study. It has been suggested that blood lead levels reflect not only acute external lead exposure but also the internal stores of lead in the bones, which can be released into the blood. Thus, blood lead levels can be considered as a steady-state mixture from both sources.36 A previous study also suggested that the effect of the cumulative lead dose (estimated by measuring lead in the tibia) may be mediated through blood lead.16 In addition, blood lead concentrations may reflect habitual and ongoing lead exposure due to environmental factors and lifestyles. Therefore, it is plausible that blood lead levels can be associated with plasma homocysteine levels 4–6 years later, as observed in the present study.
Three SNPs in the TF locus showed interactions with lead levels in the current study. TF is a monomeric glycoprotein that plays a key role in iron distribution throughout the body by transporting ferric iron among absorption, storage and utilisation sites.37 Interactions between lead exposure and the TF gene, possibly resulting from changes in iron distribution and oxidative stress status, have been reported previously. Genetic polymorphisms in the TF gene have been reported to modify the association of lead exposure with prolonged QT intervals, which serve as an indicator of electrocardiographic conduction30 and reduced intelligence quotients.31 Although three SNPs which showed interactions with lead levels are located in the non-functional elements of the TF gene, rs8649, which is in the functional region of the TF gene, is in complete linkage with rs2715632, rs2715631 and rs2715627 (|D′|=1, r2=1). Because previous reports on rs8649 are rare,38 further studies are necessary to thoroughly understand the underlying biological mechanisms of the present results.
Positive associations between lead and homocysteine were only observed in participants with variant alleles in the selected SNPs of the TF gene, whereas homocysteine levels were lower in general in these groups (see table S4 in the online supplementary file 1). Genetic polymorphisms in the TF gene can induce changes in iron distribution, haeme synthesis and activity of CBS, a haeme-containing enzyme that metabolises homocysteine into cystathionine in a transsulfuration pathway, resulting in a gradient in homocysteine levels.39–41 Although lead may also decrease haeme and haemoglobin synthesis and CBS activity, additional synergistic increases associated with lead exposure may not occur at high levels of homocysteine, possibly because of the already-low CBS activity.19 However, mechanistic evidence directly supporting the present observation is limited, and further studies focusing on the suggested pathway are warranted.
In this study, several genetic polymorphisms in the MTR and MTHFR genes, which did not show interactions with lead levels in the 1-df interaction test, were screened in the 2-df joint test. Because the 2-df joint test assesses the main genetic effects and gene–environment interactions simultaneously,23 the present results may have been driven by the association between genetic polymorphisms in the MTR and MTHFR genes and homocysteine levels, rather than interactions with blood lead levels (table 3; table S5 in the online supplementary file 1). The association between SNPs in the MTR and MTHFR genes and homocysteine levels has also been reported previously.24 42–44 Although we did not find associations of variants in the HFE, CBS and BHMT genes with homocysteine levels or interactions with lead levels, the present results should be interpreted cautiously because of the small sample size and restricted number of SNPs included in the analyses.
We assessed previously suggested biological pathways underlying the association between lead exposure and homocysteine levels by conducting a gene–environment interaction study; such studies are rarely performed but they help to strengthen the causality of observed associations by providing insights into the underlying mechanisms. However, the present study also has several limitations. First, the SNP genotyping chip used in the present study was originally developed to identify potential candidate genes in the genome-wide association study, rather than to evaluate specific candidate genes. Although we selected genes based on assumed pathways and conducted a candidate-gene approach analysis, our results should be confirmed in further studies using an independent data set to reduce the possibility of false-positive findings. In addition, functional studies are warranted to investigate the expression of the identified genes and confirm the suggested biological pathways. Second, since the Bonferroni correction was only applied in the second step, some false-positive findings may be present in the first step. We assume that the possibility of false positives is substantially reduced in the final results with the Bonferroni correction; however, methodological improvements are necessary to address issues related to lower statistical power while restricting the false-positive findings in the gene–environment interaction study. Third, subgroup analysis could not be conducted in some instances—such as for genotype—because of the small sample size (n=386). Fourth, blood lead concentrations measured only once were used as our exposure indicator. Because homocysteine levels were measured 4–6 years later, the cortical bone lead concentration—which estimates cumulative lead exposure—may have been a more suitable measure. Although blood lead levels are not only an indicator of acute lead exposure but also reflect both acute external exposure and internal stores in bones and lifestyle factors did not change substantially during the study period in the present study, time gap between lead and homocysteine measurements may be a potential source of misclassification and confounding. Therefore, further studies employing repeated measurements of blood lead levels and complementary measurements of bone lead levels are warranted to confirm the results of the present study. Fifth, SNPs previously reported to show interactions with lead exposure or to be associated with homocysteine levels, such as rs1049296 in the TF gene29–31 and rs1801133 in the MTHFR gene24 43 45 were not genotyped. However, in the present study, rs1049296 in the TF gene was imputed and assessed. Although rs1801133 in the MTHFR gene was not itself imputed, SNPs such as rs4846052 that are in the same linkage disequilibrium block and in complete linkage with rs1801133 (|D′|=1) were imputed and evaluated.
Conclusions
We found an association between blood lead levels and plasma homocysteine levels using a well-designed community-based cohort. The TF gene was found to modify the association between lead and homocysteine levels, suggesting that iron distribution may play a role in the effect of lead exposure on homocysteine levels by affecting haeme synthesis and CBS activity. However, these results should be interpreted cautiously and should be confirmed in further studies. The current findings emphasise the importance of continuous efforts to reduce human lead exposure, even in countries where lead levels are already considered low. In addition, the present results suggest a direction for future mechanistic studies.
Acknowledgments
The authors thank the study participants, research workers and technicians of the KoGES.
References
Footnotes
Contributors All authors equally contributed in designing the study. KNK and YCH analysed the data and wrote the manuscript. MRL and YHL revised the manuscript. All authors approved the version to be published.
Funding This study was supported by grants from the Korea Centers for Disease Control, Republic of Korea (2001-347-6111-221, 2002-347-6111-221, 2003-347-6111-221, 2004-E71001-00, 2005-E71001-00, 2006-E71005-00, 2007-E71001-00 and 2008-E71001-00). This study was also supported in part by the R&D Program for Society of the National Research Foundation funded by the Ministry of Science, ICT & Future Planning, Republic of Korea (2014M3C8A5030619). The funding bodies had no role in the study design, data collection, analysis, interpretation of results and manuscript writing.
Competing interests None declared.
Ethics approval The Institutional Review Board of Seoul National University Hospital approved the study protocol (C-1306-046-495).
Provenance and peer review Not commissioned; externally peer reviewed.